
研究概要
概要
私たちの研究室では、網膜疾患治療薬の効率的な薬物送達を期して、循環血液から網膜への薬物や栄養物の輸送機構に主眼を置いて研究を展開しています。網膜は視覚を司る重要な神経組織であり、後眼部の眼球壁において最も内側に存在する薄膜(約0.5 mm)です。糖尿病網膜症や網膜色素変性症、加齢黄斑変性症などの網膜疾患は、いずれも失明に繋がる重篤な網膜疾患として知られ、その治療法の向上は高齢化が進行する我が国に限らず、世界的にも重要な課題と考えられます。しかし、今なお、網膜疾患薬物療法は眼科領域の弱点の1つとされ、その最大の要因は網膜への薬物送達が極めて困難な点が挙げられます。眼への薬物投与方法と言えば、点眼が広く認識されていますが、点眼された薬物は、涙液での希釈やその他のバリア機構によってほとんど網膜に到達できません。
網膜には、循環血液と網膜神経組織を隔てる血液網膜関門(blood-retinal barrier: BRB)と呼ばれる関門機構が存在し、網膜毛細血管内皮細胞による内側血液網膜関門(inner blood-retinal barrier: inner BRB)と網膜色素上皮細胞による外側血液網膜関門(outer BRB)によって、網膜への物質移行性は厳密に制限されています。関門機構は不要な物質は通さず、必要な物質のみを細胞や組織内部へ取り入れる、いわば関所のような働きを有しており、これを利用した薬物送達法の開発に向けて、この関門メカニズムの解明にチャレンジしています。
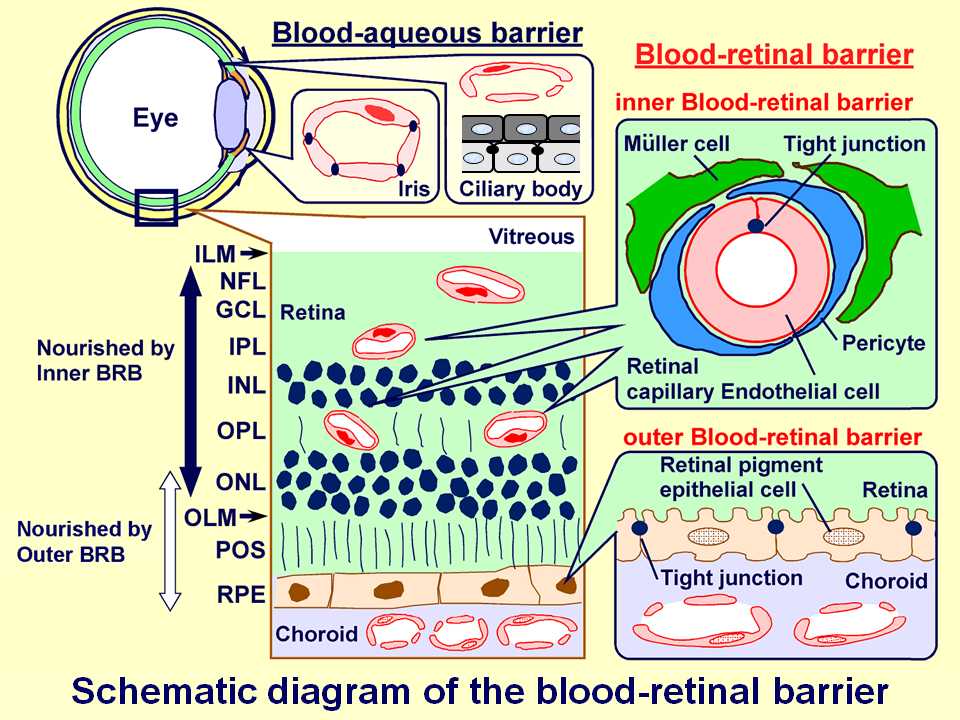
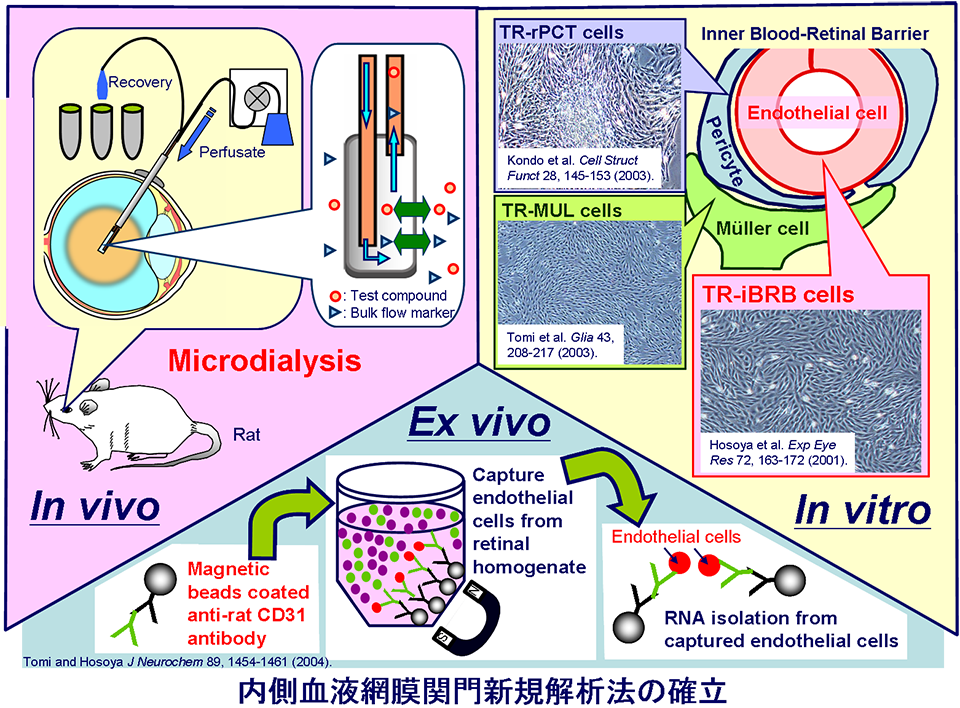
BRB研究のための解析法の確立
網膜や脳に関門が存在することは、100年以上前から知られており、脳の血液脳関門(blood-brain barrier: BBB)に関しては、長い間、大変盛んな研究が展開されてきました。微小組織である網膜に関しては、その関門機能の解明がほとんど進んでおらず、かつて、重要な研究ツールであったウシ初代培養網膜毛細血管内皮細胞を用いた解析も限界点に達していました。
私たちは、『血液網膜関門の輸送機構解析には、繁用可能なモデル細胞解析系が必須である。』との認識の下、温度感受性SV40T抗原遺伝子導入トランスジェニックラットを用いて、網膜関門細胞株の樹立に成功しました。この中には、網膜毛細血管内皮細胞株(TR-iBRB細胞)やペリサイト株(TR-rPCT細胞)、Muller細胞株(TR-MUL細胞)などがあり、これら細胞株を用いることでBRB機能の包括的解明が可能となりました。以上に加え、私たちは、磁気標識抗CD31抗体を用いた内皮細胞高純度単離法を利用した血液網膜関門遺伝子発現定量法の確立や、ラットの硝子体内に薬物を捕獲するプローブ(microdialysis)を挿入し、眼球内濃度を経時的に測定することによって網膜からの排出輸送を解析する手法の確立にも成功しています。
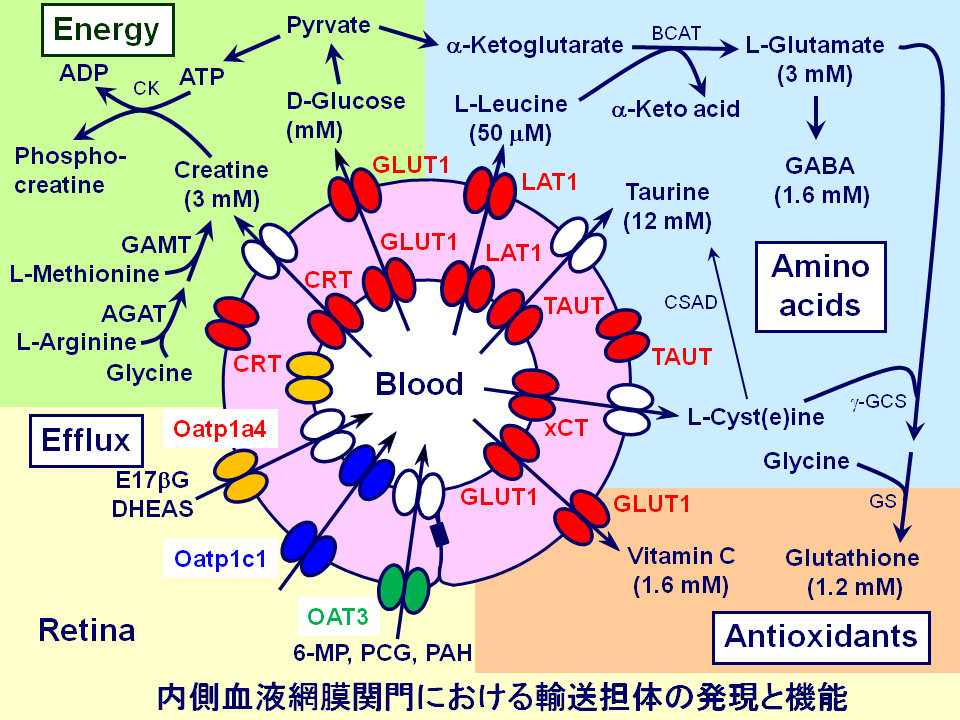
血液網膜関門を介した栄養物供給機構
網膜は光を受容する重要な神経組織であり、また、酸化などのストレスに曝されている組織でもあります。このため、健全な視覚機能の維持において、網膜への栄養物供給は重要な生理機構の1つと考えられます。
私たちは、網膜毛細血管内皮細胞株(TR-iBRB細胞)や遺伝子発現定量法、生理条件下での血液網膜関門輸送機構解析法を駆使して、血液網膜関門を介した神経伝達物質の前駆物質や抗酸化作用物質などの供給機構を検証してきました。
ラットにおける網膜取り込み速度やTR-iBRB細胞を用いた輸送機能解析の結果、循環血液から網膜へのビタミンC供給は、酸化型アスコルビン酸であるデヒドロアスコルビン酸を輸送するグルコース輸送担体GLUTsを介することが示唆されました。また、発現解析では、ラット網膜血管内皮細胞におけるGLUT1の高発現が示され、網膜へのビタミンC供給がGLUT1を介することが示唆されました。網膜において、タンパク質合成に不可欠なアミノ酸であると同時に、神経伝達物質であるグルタミン酸の主要な前駆物質となるL-ロイシンの供給に関しては、ラットにおける網膜取り込み速度やTR-iBRB細胞を用いた輸送機能解析によって、inner BRBに発現する中性アミノ酸トランスポーター・LAT1の寄与が示唆されました。LAT1は、アミノ酸に類似する薬物を輸送することでも知られており、網膜への薬物送達での利用が期待されます。私たちのこれまでの研究によって、網膜の生理機能に重要な栄養物であるビオチンやアセチル-L-カルニチン、グリシン、葉酸、タウリン、ニコチン酸、L-アルギニン、L-セリン、L-シスチン、アデノシンなどの網膜への供給に、それぞれSMVT、OCTN2、GlyT1、RFC1、TAUT、MCTs、CAT1、ASCTs、xCT、ENT2などのトランスポーターの関与が明らかとなりました。近年では、網膜のクレアチン恒常性維持機構や、網膜へのコリンやL-カルノシン供給機構、網膜グリア細胞であるMuller細胞におけるアデノシンとヒポキサンチン濃度制御機構などに関する研究成果を発表しています。
血液網膜関門を介したアニオン性化合物輸送機構
網膜内の神経伝達物質や各種ステロイドホルモンは、代謝によって不活性化されることが知られていましたが、その後の除去機構の詳細は明らかとされていませんでした。私たちは、BRBの重要な役割の1つとして、「網膜内で産生される不要物質の除去」を提案し、その実証にチャレンジしてきました。私たちは、microdialysisを駆使して、ステロイドホルモン代謝物の1つであるエストラジオールグルクロン酸抱合体の網膜からの消失を解析し、この機構に有機アニオントランスポーターの関与を見出しました。阻害解析や高純度網膜毛細血管内皮細胞画分を用いた発現解析の結果、網膜からのエストラジオールグルクロン酸抱合体の除去に、Oatp1a4の寄与が示唆されました。このことは、免疫組織化学的解析において、Oatp1a4タンパク質がinner BRBの網膜側細胞膜と血液側細胞膜に局在していたことからも支持されます。また、私たちのこれまでの解析結果から、有機アニオントランスポーター・OAT3もBRBを介した網膜からのアニオン性化合物排出輸送に寄与することが示唆されています。免疫組織化学的解析では、OAT3タンパク質がinner BRBの網膜側細胞膜に局在することが示されました。OAT3は抗生物質であるベンジルペニシリンや白血病治療薬である6-メルカプトプリンを基質とすることが知られており、microdialysis法を用いた私たちの解析では、これら薬物の網膜移行がBRBに発現するOAT3によって積極的に妨げられていることが示唆されました。網膜への薬物送達を実現する上で、Oatp1a4やOAT3の機能を組み込んだ評価が重要であると考えられます。
血液網膜関門における輸送の制御機構
重篤な網膜疾患が発症した場合、網膜内では、酸化や虚血、飢餓、浸透圧変動など多様なストレスが惹起され、これに応答して、BRBの輸送機構が変動することが想定されます。このような病態時にこそ、網膜疾患治療薬が投与されることから、BRBの輸送機構の病態時変動やストレス応答性を明らかにすることは、網膜疾患治療の向上に重要な視点と考えられます。
私たちは、これまでに、inner BRBに発現するL-シスチントランスポーター・xCTの発現量が、酸化ストレスに応答して変動することを見出し、この発現制御機構によって、抗酸化物質であるグルタチオンの合成が制御されることに寄与することを示唆しました。さらに、近年では、inner BRBの中性アミノ酸トランスポーター・LAT1の発現がグルコース欠乏に応答して変動することや、Muller細胞に発現するTAUTの発現量が浸透圧変化に応答して変動することを報告しています。
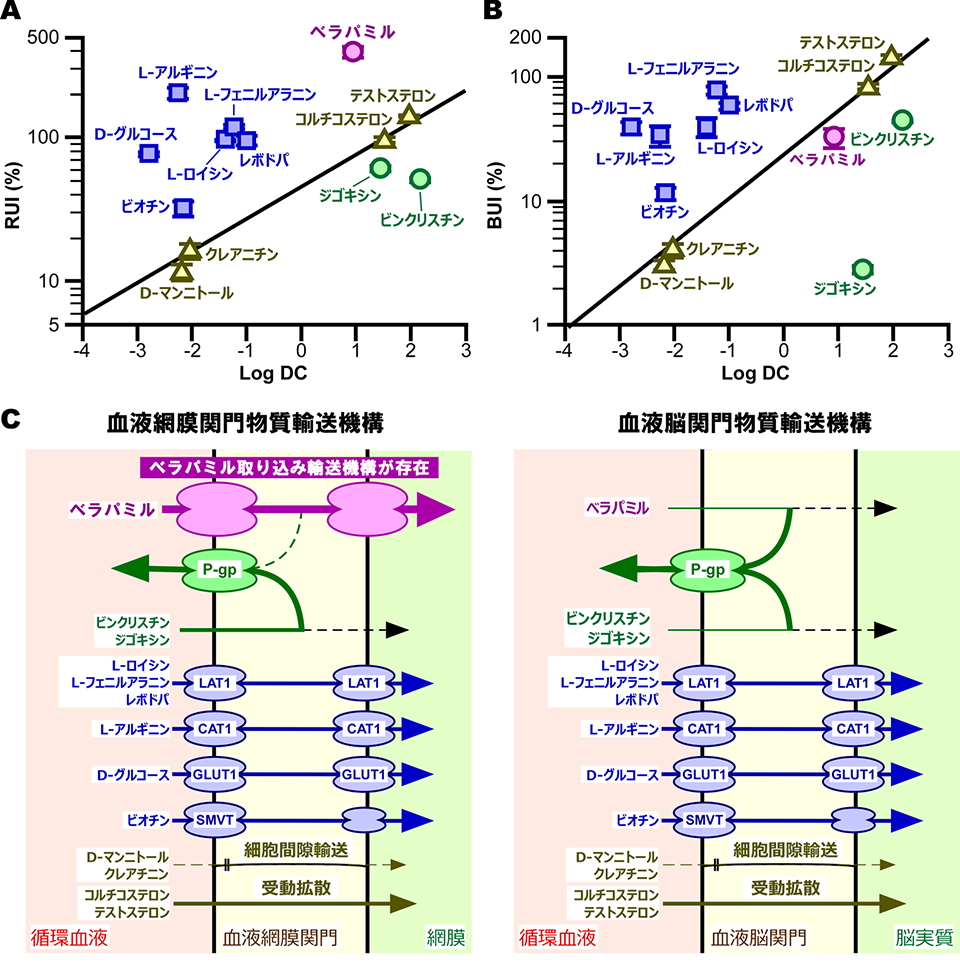
BRBとBBBの異なる関門特性
製薬産業で展開される効率的な新薬開発においては、簡便で迅速な候補化合物の探索法が必須と考えられます。私たちは、薬物や栄養物のBRBを介した網膜移行性に関してin vitro-in vivo相関解析を実施し、TR-iBRB細胞を用いたin vitro輸送解析が化合物の網膜移行性予測に有用性であることを報告しました。
In vitro-in vivo相関解析は、BRBを受動拡散によって透過する化合物群(グループA)と担体介在輸送によって透過する化合物群(グループB)に分類して実施し、グループAが脂溶性-RUI値間での相関性(相関式1)を示した一方、グループBはいかなる相関性も示しませんでした。また、化合物の血液脳関門(BBB)透過性評価に有用なbrain uptake index (BUI)に関しても相関解析を実施し、グループAのみが相関性を示す結果となりました。D-グルコースやアミノ酸、レボドパ、ビオチンのRUI値とBUI値はともに、相関式から予測されるよりも高い値を示し、両関門では、GLUT1やCAT1,LAT1,SMVTなどのトランスポーターが同様に寄与することが示唆されました。一方、興味深いことに、カチオン性薬物であるベラパミルは相関式2と同程度のBUI値を示す一方、そのRUI値は相関式1よりも高い結果となりました。ベラパミルやビンクリスチン、ジゴキシンは排出トランスポーター・P-gpの基質であり、これら薬物の低い脳移行性は有名です。BRBでのP-gp発現も報告されており、このことは、ビンクリスチンなどのRUI値からも支持されます。以上から、inner BRBとBBBはカチオン性薬物に対して異なる関門特性を有することが示され、inner BRBにおけるカチオン性薬物取り込み輸送機構の存在が示唆されました。
相関式1; RUI = 46.2 × exp(0.515 × log DC) (r2 = 0.807, p < 0.01)
相関式2; BUI = 24.2 × exp(0.816 × log DC) (r2 = 0.885, p < 0.01)
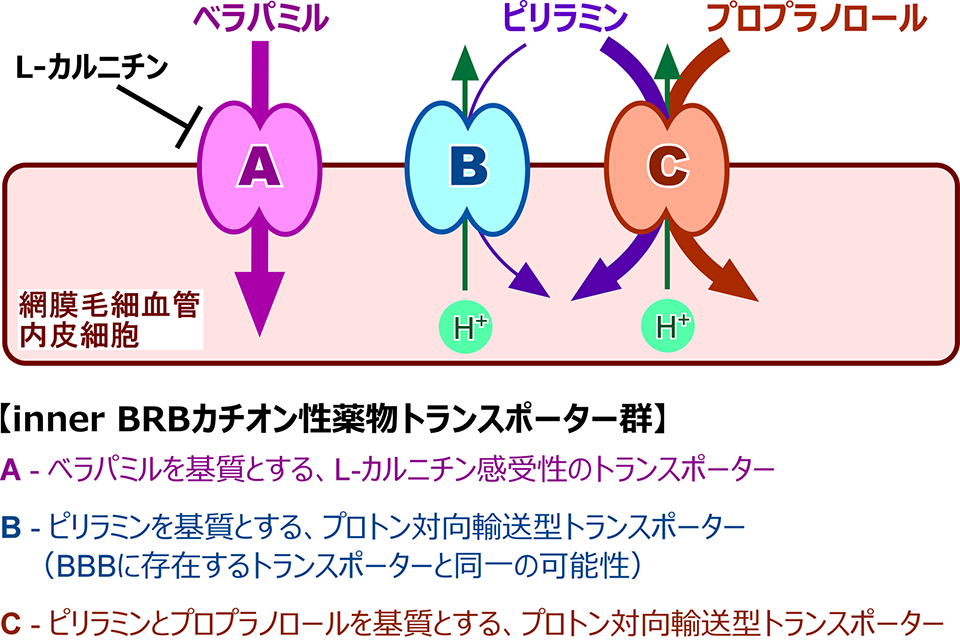
血液網膜関門を介したカチオン性薬物輸送機構
近年、神経保護作用を有する薬物が脳や網膜の神経疾患治療に有用であることや、プロプラノロールが未熟児網膜症治療に有用であることなどが報告されています。このような有益な薬物には、多数のカチオン性薬物が含まれており、BRBを介したカチオン性薬物取り込み輸送機構の検証は、網膜疾患治療法の向上に寄与するものと期待されます。私たちは、ベラパミルやプロプラノロール、ピリラミンなどのカチオン性薬物に関して、取り込み輸送機構を解析した結果、異なる複数のカチオン性薬物輸送系がinner BRBに存在することが明らかとなりました。また、神経保護作用を有するデシプラミンやメマンチンなどが、これら輸送系を阻害することから、カチオン性薬物取り込み輸送機構を介した網膜疾患治療薬の効率的送達が期待されます。
Inner BRBのカチオン性薬物輸送機構はキニジンやアマンタジン、クロニジンなどのカチオン性薬物によって阻害されることから、その分子実体が有機カチオントランスポーターであると推測されました。しかし、inner BRBのカチオン性薬物輸送機構は、既知の有機カチオントランスポーターの基質や阻害剤に対して非感受性であり、inner BRBのカチオン性薬物取り込み輸送機構は未知の有機カチオントランスポーターによって担われると考えられます。近年、未知分子によるカチオン性薬物輸送は他の組織においても見出されており、従来とは性質を異にする新規格トランスポーター分子の解明は、網膜や脳への薬物移行性予測に向けた新しいフロンティアと考えています。
血液脳関門を介したニコチン取り込み輸送機構
ニコチンはタバコに含まれる有害物質として広く知られていますが、循環血液から脳への移行機構に関しては、あまりよく知られていませんでした。私たちは、ラットにおけるin vivo取り込み輸送解析や脳毛細血管内皮細胞株を用いたin vitro輸送解析を実施し、血液脳関門(BBB)にニコチン(タバコ成分)取り込み輸送機構が存在すること、また、その輸送機構が担体介在輸送であることを明らかとしました。ニコチンは生理条件下においてカチオン性を示すことが知られており、BBBを介したニコチン輸送機構の分子実体として、何かしらの有機カチオントランスポーターの関与が考えられます。以上の知見は、喫煙などの際に、ニコチンが脳に及ぼす影響を詳しく理解する上で重要なものと考えられます。
加えて、近年、ニコチンやその誘導体は、ニコチン性アセチルコリン受容体(nAChR)を薬理ターゲットとする中枢疾患治療薬の有力候補化合物と目されています。BBBを介したニコチン輸送機構に限らず、さまざまな臓器や組織に存在するニコチン移行性制御機構を明らかとすることは、アルツハイマー病などの重篤な中枢疾患治療の向上に寄与するものと期待されます。
脳内生理活性物質恒常性維持への脳関門の役割
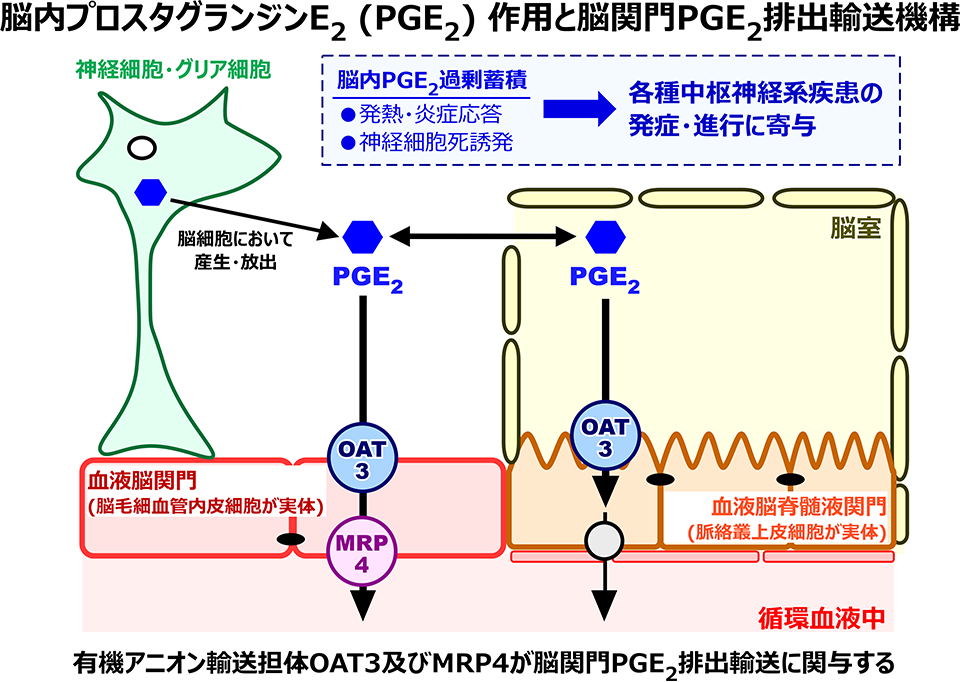
中枢神経系と循環血液とを隔てる脳関門は、脳-循環血液間の物質交換を制御しています。脳関門は、循環血液と脳実質とを隔てる血液脳関門 (BBB) と、循環血液と脳脊髄液 (CSF) とを隔てる血液脳脊髄液関門 (blood-CSF barrier: BCSFB) によって構成されています。脳への薬物送達を考える上で、これら関門組織の重要性は言うまでもありませんが、これに加えて、近年では、脳内不要物質の除去にBBBやBCSFBの排出輸送機構の関与が明らかとなってきました。不要物質の脳内過剰蓄積は中枢神経系疾患時に認められ、病態進行に関与することが知られていることから、脳内不要物質の除去機構解明は、脳内環境正常化を目的とした創薬に貢献するものと期待されます。これまでに私たちは、各種in vivo/in vitro脳関門排出輸送解析法を駆使し、てんかんやアルツハイマー病時での脳内蓄積が知られるプロスタグランジンE2 (PGE2) およびグアニジノ化合物の脳からの除去に脳関門が関与することを明らかとしました。グアニジノ化合物であるグアニジノ酢酸やクレアチニンに関しては、in vivo解析によって、BCSFBを介した促進的排出輸送を実証しました。In vivo遺伝子ノックダウン法による解析では、BCSFBを介したグアニジノ酢酸排出輸送は主にCRTが、クレアチニン排出輸送はCRTと有機カチオントランスポーター・OCT3が担うことを明らかにしました。PGE2はBBBを介し排出輸送され、その過程にBBB脳側膜に局在するトランスポーター・OAT3とBBB血液側膜に局在するトランスポーター・MRP4の関与を明らかとしました。また、BCSFBを介したPGE2排出輸送も存在し、これをOAT3とプロスタグランジントランスポーター (PGT) が担うことを見出しました。さらに、私たちは、炎症惹起下や過剰L-グルタミン酸共存下に、BBBを介したPGE2排出輸送が減弱することを見出しました。脳炎症や興奮性神経伝達過剰な疾患では、脳内PGE2濃度上昇が知られており、これら疾患の発症・進行にBBBを介したPGE2輸送系の関与が考えられます。脳関門のPGE2排出輸送機能の正常化は中枢神経系疾患治療戦略に直結するものと期待されることから、私たちは、PGE2排出機能減弱に関する詳細なメカニズムの解明に挑んでいます。
展望
血液網膜関門の輸送機序の研究は緒に就いたばかりであり、世界的にみても研究グループはそう多くはありません。一方、高齢化社会を迎え、糖尿病網膜症、加齢黄斑変性症や緑内障による網膜変性症などの網膜疾患に対する薬物治療法開発の重要性は益々増大しています。点眼による網膜への薬物送達はほぼ不可能であるため、血液から血液網膜関門を介した網膜への薬物送達による治療法が望まれており、「飲む目薬」開発を目指しております。血液網膜関門の輸送機序解明を通し、医療のより良い発展の一助となることを願っています。
