








| 連絡先 |
| 〒930-0194 富山市杉谷2630 富山大学学術研究部薬学・和漢系 生体界面化学研究室 TEL: 076-434-7565 FAX: 076-434-7568 |


生体界面化学研究室は、その名の如く、界面化学をベースにした研究手法を用いて生体(特に生体膜上)での現象を解明することと、コロイド化学的知見を活かして製剤開発に寄与することを目標にしています。
生体膜は細胞や細胞内小器官の内と外を分ける境界(界面, interface)の役割を果たしており、エネルギー変換や物質輸送、情報伝達といった多彩な生命活動の舞台となっています。生物は自身と外界とを区別 する境界膜を、脂質分子の自己組織化によって獲得し、剛直な膜では不可能な物理的柔軟性と物質選択的透過性を達成している訳です。数千もの脂質種が生体内 に存在するという事実は明らかに、各脂質固有の生化学的特性が生体に高度な機能を与えていることを示していますが、これらの特性が発揮される場がやはり生 体膜上であることを考えれば、全ての生命活動が生体膜の分子集合体としての物理化学的特性に裏打ちされたものであることを忘れてはなりません。
このような観点から、我々は、生物物理学、生体界面化学の立場から、生体膜での生命現象を、空間構造(Å-μm)と時間構造(ns-min)の両面から明 らかにする研究を展開しています。生体膜を構成している主な成分であるリン脂質分子は、自発的に集合して脂質二重膜を形成します。この膜は静止して変化し ないものではなく、構成している分子が絶えず熱運動することで時間的・空間的に揺らいでいる動的で不均一な構造です。さらに、分子の代謝や輸送によって、 膜の組成や分子局在も刻々と変化しています。この代謝や輸送には、生体膜のもう一つの重要な構成成分であるタンパク質が深く関与しています。我々は、この ような脂質とタンパク質が織り成す生体界面の複雑な時空間構造を計測し、生体膜が如何にして生命機能を担っているのかを、熱・統計力学などの物理化学やコ ロイド化学の言葉を使って説明することを目指しています。また、生体膜現象の基礎科学的な側面だけでなく、脂質やタンパク質分子がつくる複合体微粒子の解 析を通じて、これを製剤へ応用することを指向した研究もおこなっています。
1.膜脂質の膜間移動ダイナミクス
生体内において、脂質の多くは小胞体で合成された後、細胞膜や各オルガネラ膜へと分配 されます。また、肝臓や小腸から分泌された脂質はリポタンパク質として血中を循環し、末梢細胞へ供給されます。このように細胞内、細胞間の脂質輸送は、生 体の恒常性維持において重要な役割を担っています。しかしながら、脂質のダイナミクスについての定量的な議論は細胞系のみならず無細胞系でもなかなか行わ れてきませんでした。それは、合理的なダイナミクス計測法が無かったからです。我々は、リン脂質ベシクルを用いて、脂質の粒子間移動を、蛍光エネルギー移 動(FRET)や分子間エキシマー蛍光を利用した手法や、中性子散乱法を使って速度論的に解析しています。現在は脂質輸送活性をもつタンパク質のアッセイ 系としての応用を図っています。
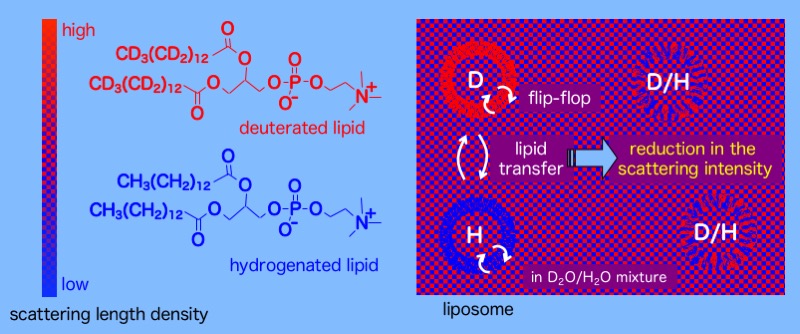
中性子散乱によるリン脂質のダイナミクスの評価法
2.膜脂質のフリップフロップ
細胞膜(形質膜)では脂質二重層の細胞質側と外側とで脂質組成が大きく異なっており、アポトーシスが誘導されると、非対称性が崩れ、細胞質側に局在してい たホスファチジルセリンが外側に露出し、これがマクロファージによる貪食のシグナルとなります。すなわち細胞膜の非対称性の破綻は細胞死に直結します。一 方、脂質生合成の場である小胞体膜では、新生されたリン脂質は脂質二重層の細胞質側に導入された後、二重層の内側と外側の脂質数のバランスを保つために速 やかに内腔側に移行(フリップ)されます。このように脂質のダイナミクスを制御することは生命活動において必要不可欠な問題です。我々は、リン脂質のフ リップフロップ速度を、蛍光消光法や中性子散乱法を用いて評価し、フリップフロップの促進に関わる因子を明らかにする研究を進めています。また、人工的に ベシクル膜に非対称性を付与する系も構築しています。
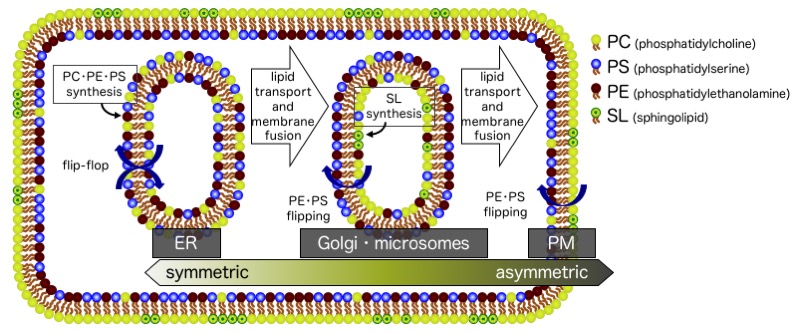
生体膜リン脂質の対称・非対称分布
3.ナノディスク
高密度リポタンパク質(HDL)は末梢のコレステロールを肝臓へ輸送するコレステロール逆転送系を担う血中コロイド粒子です。この粒子は肝臓や末梢細胞で 生成されますが、生成時にはディスク状を呈しています。このディスク状粒子は、リン脂質とHDLの構成タンパク質であるアポリポタンパク質A-I (apoA-I)あるいは類似の構造をもつ両親媒性ペプチドを用いて人工的に作成することが可能で、ナノディスクとよばれています。我々は様々な蛍光解析 法を駆使して、ナノディスクの構造解明を行っています。蛍光寿命や分子内エキシマー蛍光を利用した評価法により、リン脂質含有量の少ない小さなナノディス クは、脂質二重層が平板状ではなく鞍状曲面を形成していることを突き止めました。また、SANS法による解析の結果、リン脂質のダイナミクスがLUVとは 大きく異なり、ナノディスクからのリン脂質やコレステロールの解離速度がLUVからの解離よりも著しく速いことを明らかにしています。また両親媒性ペプチ ドを使って、ナノディスクが自己複製される系や、ペプチドのライゲーションによって構造を安定化させる系を構築しています。ナノディスクは生理的に重要で あるばかりでなく、膜タンパク質の再構成ツールや、ドラッグデリバリーシステムへの応用が行われており、ナノディスクの物理化学的特性を明らかにする意義 は非常に大きいのです。
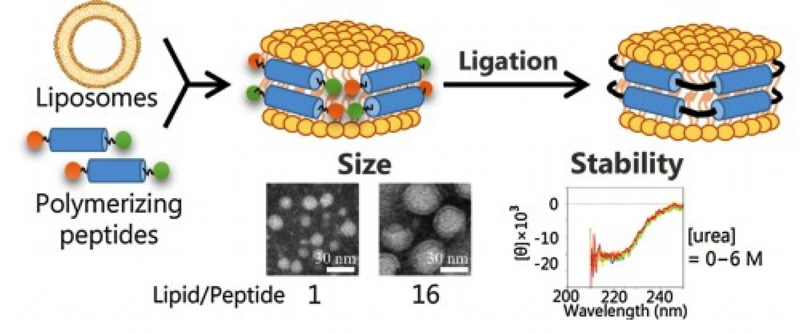
ペプチドのライゲーションにより構造を安定化させたナノディスク (Colloid Surf. B 146, 423 (2016))
4.膜の曲率とタンパク質の相互作用
生体膜は膜融合などの過程で膜の曲率を大きく変遷させます。また、膜の曲率を誘起したり、曲率を認識したりするタンパク質が多数見つかっています。我々 は、曲率が膜環境をどのように変え、それがタンパク質との相互作用にどのように影響を与えるのかを、曲率の異なるベシクルを使って評価しています。膜の曲 率がエンタルピーを増加させることを等温滴定型カロリメトリーを使った手法によって明らかにしました。また、アミロイドβタンパク質の膜への結合とアミロ イド繊維化は高曲率膜において促進されることを見いだしました。膜曲率変化の生理的意義の解明を目指した研究を進めています。
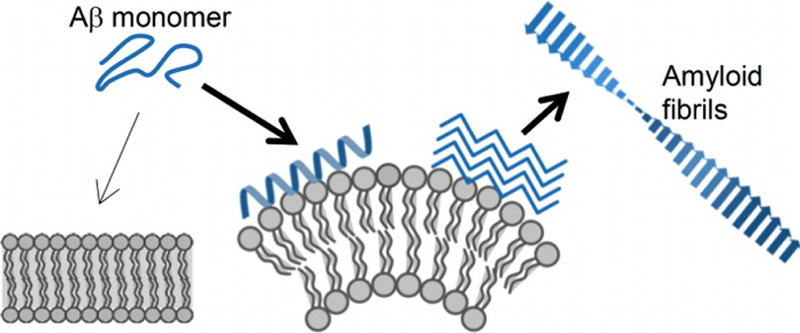
高曲率膜はアミロイドβタンパク質の結合を高め、繊維化を促進する (Langmuir 31, 11549 (2015))
5.非ラメラ相形成脂質と新規脂質微粒子
生体膜は二分子膜構造を形成していますが、その中には非ラメラ相形成脂質と呼ばれる、負の自発曲率を持つ脂質が多数存在しています。このような脂質は膜中 の側方圧を変化させ、タンパク質の膜への結合性や、膜タンパク質の構造や活性を調節すると考えられます。蛍光法やNMRによって膜の側方圧変化を検出する 手法を考案し、非ラメラ相形成脂質の効果を明らかにしてきました。一方、これらの脂質が形成する非ラメラ相(逆ヘキサゴナル相、キュービック相)は水中で の分散安定化が困難とされてきましたが、高分子乳化剤を用いた方法により100-200 nmの新規コロイド粒子(キューボソーム、ヘキソゾーム)の調製法、評価法を確立しました。最近ではさらに、脂質や乳化剤に工夫を施すことで、液晶構造 や、血漿タンパクとの相互作用を調節できることも分かってきました。このように、逆ヘキサゴナル相やキュービック相の物性と生物物理学的機能の解明が着々 と進んでおり、その医薬品製剤応用への道も開かれつつあります。
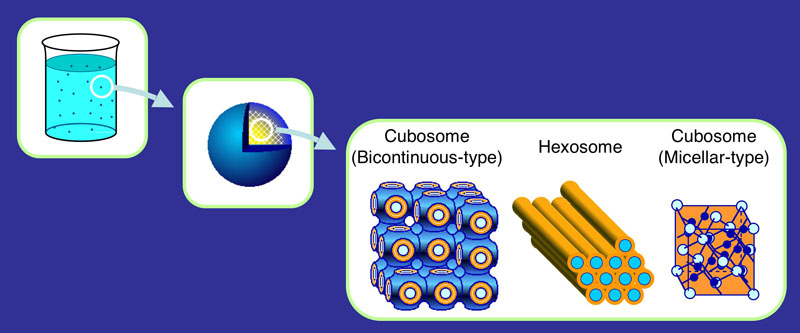
キューボソーム、ヘキソゾームの模式図